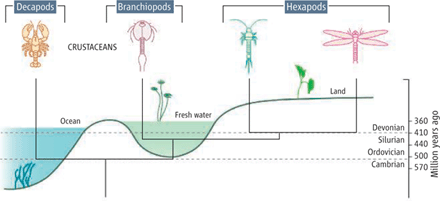
Hexapod evolution. The last common ancestor of hexapods and crustaceans (branchiopods, specifically) may have originated in freshwater during the Late Silurian, giving rise to extant freshwater dwelling branchiopods (fairy shrimps, water fleas, and tadpole shrimps) and insects. This hypothesis accounts for the missing fossil record of branchiopods and hexapods before the Devonian. (Fig. 1, Glenner, et al., Science 22 December 2006: Vol. 314. no. 5807, pp. 1883 - 1884)
I've always been fascinated by the subject of insect evolution. People with even a rudimentary appreciation for detail will have noted some of the similarities between certain insects and crustaceans -- the sort of exoskeletons and segmentation they possess, for one thing. However, although scientists have known that there is shared ancestry between the groups, hard transitional fossils for insects have never been found. Now, new morphological and genetic research (Hox) indicates that insects may not be cousins with crustaceans, sharing a common ancestor, but may have actually originated within the clade, most closely related to branchiopods. Below the fold, I am pasting in the full-text of a recent Science article (Glenner, et al., Science 22 December 2006: Vol. 314. no. 5807, pp. 1883 - 1884) giving perspective on six papers detailing the genomic relationships among these groups:
__________
Science 22 December 2006:
Vol. 314. no. 5807, pp. 1883 - 1884
DOI: 10.1126/science.1129844
Perspectives
EVOLUTION:
The Origin of Insects
Henrik Glenner, Philip Francis Thomsen, Martin Bay Hebsgaard, Martin Vinther Sørensen, Eske Willerslev
Although hexapods--those arthropods having six legs, including insects--are the most diverse group of contemporary animals in terms of biological niches and number of species, their origin is highly debated. A key problem is the almost complete absence of fossils that connect hexapods to the other major arthropod subphyla, namely Crustacea, Myriapoda (such as centipedes and millipedes), and Chelicerata (such as scorpions and spiders). Over the years, hexapods (insects, springtails, proturnas, and diplurans) have been phylogenetically linked to all of these major arthropod taxa (1). [see Fig. 1 at top of post]
Traditionally, hexapods and the multi-legged myriapods have been united in a group named Atelocerata on the basis of morphological similarities between their tracheal respiration systems and head appendages. However, recent evidence from phylogenetic analyses of molecular sequence data from a variety of genes, as well as from newer morphological studies, points to a relationship between hexapods and crustaceans (2-9), a grouping commonly referred to as Pancrustacea. Furthermore, studies on neurological development in the major arthropod groups have pointed out similarities between the myriapods and chelicerates (10). Hence, pancrustacean monophyly seems to be gaining more support. So, what does this view tell us about the possible origin of hexapods?
The crustaceans are recorded at least as far back as the Upper Cambrian, about 511 million years ago (11), where they are found in marine sediments (see the figure). However, except for the debated Devonohexapodus bocksbergensis specimen (12, 13), all hexapod remains are found only in freshwater or terrestrial strata no earlier than the Devonian, around 410 million years ago (14). This leaves a gap of 100 million years to the earliest crustaceans. The common explanation has been that earlier traces of hexapods have been erased from the fossil record and that hexapods, like other major groups of terrestrial animals, have closely related ancestors to be found in the marine environment.
The recent morphological and molecular-based studies suggest an alternative interpretation--that hexapods originated within the crustaceans rather than as a sister group (15-20). Although the morphological studies mainly favor a close phylogenetic connection between hexapods and malacostracan crustaceans (crabs and crayfish) (15, 16), recent molecular sequence data suggest that hexapods are closely related to branchiopods (17, 19, 20), a freshwater dwelling group of crustaceans that includes water fleas and fairy shrimp. This hypothesis is supported by analysis of Hox genes that demonstrates homology between development of the pregenital trunk region in insects and the thorax in branchiopods (21). The new molecular results correspond well with the fossil record and suggest an evolutionary origin of the hexapods in freshwater around 410 million years ago rather than in the marine Cambrian environment (17).
The vast majority of extant branchiopods are freshwater animals, and the few that are found in saltwater are believed to have invaded the sea secondarily. From the fossil record, it is known that modern branchiopods date back to the Early Devonian, by which time they were fully adapted to freshwater habitats (22). This late appearance of the freshwater branchiopods corresponds exactly with the emergence of hexapods and suggests that their last common ancestor swam around in a freshwater pond sometime in the Late Silurian (423 to 416 million years ago) or Early Devonian. This corresponds well with the time split between the crustacean and hexapod lineages estimated from molecular clock analyses (23). If correct, the early marine ancestor of the hexapods might have appeared more similar to Rehbachiella kinnekullensis, a close marine relative to branchiopods from Upper Cambrian (24), than to D. bocksbergensis or other hexapods.
The successful colonization of the terrestrial environment by hexapods seems to coincide with other major groups of land pioneering animals such as the chelicerates and the myriapods in the Late Silurian and the tetrapods (amphibians, reptiles, birds, and mammals) in the Late Devonian. All these events appear to have occurred through a freshwater dwelling phase in their evolutionary transition from marine to true terrestrial animals. The Devonian is believed to have been a time of severe drought, which might have forced these animals (at least hexapods and tetrapods) onto land as their freshwater habitats vanished.
It has been a puzzle as to why hexapods--in particular insects, which possess a morphology that apparently enables them to adapt to virtually all types of terrestrial environments--have not been able to diversify successfully in the marine environment. It is likewise remarkable that the crustaceans--fulfilling a biological role in the sea comparable to the insects on land--have not been able to invade land to a greater extent despite their considerable age. The recent phylogenetic analyses of molecular sequence data suggest a paradigm shift concerning the phylogenetic position of hexapods--that crustaceans successfully invaded land as insects. It is possible that when insects entered terrestrial habitats, their crustacean ancestors had already diversified in marine environments and occupied all potential niches, which could explain why insects were prevented from colonizing the sea subsequently. Most important, however, the new molecular results offer a solution to the enigma concerning the absence of marine hexapod remains in the fossil records prior to the Devonian.
References and Notes
But remember: evolution is just a fairy-tale. All this "research" is really just a vast Darwinian conspiracy. Intelligent Design Creationism explains this stuff so much better! Nonetheless, it's fascinating stuff; to think about how much sense this makes, and how well-supported the evidence is, that Devil sure is tricky, eh?
________________
Technorati tags: Evolution
Science 22 December 2006:
Vol. 314. no. 5807, pp. 1883 - 1884
DOI: 10.1126/science.1129844
Perspectives
EVOLUTION:
The Origin of Insects
Henrik Glenner, Philip Francis Thomsen, Martin Bay Hebsgaard, Martin Vinther Sørensen, Eske Willerslev
Although hexapods--those arthropods having six legs, including insects--are the most diverse group of contemporary animals in terms of biological niches and number of species, their origin is highly debated. A key problem is the almost complete absence of fossils that connect hexapods to the other major arthropod subphyla, namely Crustacea, Myriapoda (such as centipedes and millipedes), and Chelicerata (such as scorpions and spiders). Over the years, hexapods (insects, springtails, proturnas, and diplurans) have been phylogenetically linked to all of these major arthropod taxa (1). [see Fig. 1 at top of post]
Traditionally, hexapods and the multi-legged myriapods have been united in a group named Atelocerata on the basis of morphological similarities between their tracheal respiration systems and head appendages. However, recent evidence from phylogenetic analyses of molecular sequence data from a variety of genes, as well as from newer morphological studies, points to a relationship between hexapods and crustaceans (2-9), a grouping commonly referred to as Pancrustacea. Furthermore, studies on neurological development in the major arthropod groups have pointed out similarities between the myriapods and chelicerates (10). Hence, pancrustacean monophyly seems to be gaining more support. So, what does this view tell us about the possible origin of hexapods?
The crustaceans are recorded at least as far back as the Upper Cambrian, about 511 million years ago (11), where they are found in marine sediments (see the figure). However, except for the debated Devonohexapodus bocksbergensis specimen (12, 13), all hexapod remains are found only in freshwater or terrestrial strata no earlier than the Devonian, around 410 million years ago (14). This leaves a gap of 100 million years to the earliest crustaceans. The common explanation has been that earlier traces of hexapods have been erased from the fossil record and that hexapods, like other major groups of terrestrial animals, have closely related ancestors to be found in the marine environment.
The recent morphological and molecular-based studies suggest an alternative interpretation--that hexapods originated within the crustaceans rather than as a sister group (15-20). Although the morphological studies mainly favor a close phylogenetic connection between hexapods and malacostracan crustaceans (crabs and crayfish) (15, 16), recent molecular sequence data suggest that hexapods are closely related to branchiopods (17, 19, 20), a freshwater dwelling group of crustaceans that includes water fleas and fairy shrimp. This hypothesis is supported by analysis of Hox genes that demonstrates homology between development of the pregenital trunk region in insects and the thorax in branchiopods (21). The new molecular results correspond well with the fossil record and suggest an evolutionary origin of the hexapods in freshwater around 410 million years ago rather than in the marine Cambrian environment (17).
The vast majority of extant branchiopods are freshwater animals, and the few that are found in saltwater are believed to have invaded the sea secondarily. From the fossil record, it is known that modern branchiopods date back to the Early Devonian, by which time they were fully adapted to freshwater habitats (22). This late appearance of the freshwater branchiopods corresponds exactly with the emergence of hexapods and suggests that their last common ancestor swam around in a freshwater pond sometime in the Late Silurian (423 to 416 million years ago) or Early Devonian. This corresponds well with the time split between the crustacean and hexapod lineages estimated from molecular clock analyses (23). If correct, the early marine ancestor of the hexapods might have appeared more similar to Rehbachiella kinnekullensis, a close marine relative to branchiopods from Upper Cambrian (24), than to D. bocksbergensis or other hexapods.
The successful colonization of the terrestrial environment by hexapods seems to coincide with other major groups of land pioneering animals such as the chelicerates and the myriapods in the Late Silurian and the tetrapods (amphibians, reptiles, birds, and mammals) in the Late Devonian. All these events appear to have occurred through a freshwater dwelling phase in their evolutionary transition from marine to true terrestrial animals. The Devonian is believed to have been a time of severe drought, which might have forced these animals (at least hexapods and tetrapods) onto land as their freshwater habitats vanished.
It has been a puzzle as to why hexapods--in particular insects, which possess a morphology that apparently enables them to adapt to virtually all types of terrestrial environments--have not been able to diversify successfully in the marine environment. It is likewise remarkable that the crustaceans--fulfilling a biological role in the sea comparable to the insects on land--have not been able to invade land to a greater extent despite their considerable age. The recent phylogenetic analyses of molecular sequence data suggest a paradigm shift concerning the phylogenetic position of hexapods--that crustaceans successfully invaded land as insects. It is possible that when insects entered terrestrial habitats, their crustacean ancestors had already diversified in marine environments and occupied all potential niches, which could explain why insects were prevented from colonizing the sea subsequently. Most important, however, the new molecular results offer a solution to the enigma concerning the absence of marine hexapod remains in the fossil records prior to the Devonian.
References and Notes
- G. Giribet, C. Ribera, Cladistics 16, 204 (2000).
- J. M. Mallatt et al., Mol. Phy. Evol. 31, 178 (2004).
- J. C. Regier, J. W. Shultz, Mol. Phy. Evol. 20, 136 (2001).
- J. W. Shultz, J. C. Regier, Proc. R. Soc. London Ser. B 267, 1011 (2000).
- U. W. Hwang et al., Nature 413, 154 (2001).
- G. Giribet et al., Nature 413, 157 (2001).
- C. E Cook et al., Curr. Biol. 11, 759 (2001).
- Y.-x. Luan et al., Mol. Biol. Evol. 22, 1579 (2005).
- W. Dohle, Ann. Soc. Ent. France 37, 85 (2001)
- H. Dove, A. Stollewerk, Development 130, 2161 (2003).
- G. E. Budd et al., Science 294, 2047a (2001).
- F. Hass et al., Org. Divers. Evol. 3, 39 (2003).
- R. Willmann, Org. Divers. Evol. 5, 199 (2005).
- M. S. Engel, D. A. Grimaldi, Nature 427, 627 (2004).
- S. Harzsch, Integrative Comp. Biol. 46, 162 (2006).
- N. J. Strausfeld, Arthropod Struct. Dev. 34, 235 (2005).
- J. C. Regier, J. W. Shultz, Mol. Biol. Evol. 14, 902 (1997).
- C. E. Cook, Q. Y. Yue, M. Akam, Proc. R. Soc. London Ser. B 272, 1295 (2005).
- J. C. Regier et al., Proc. R. Soc. London Ser. B 272, 395 (2005).
- J. Mallatt, G. Giribet, Mol. Phy. Evol. 40, 772 (2006).
- M. Averof, M. Akam, Nature 376, 420 (1996).
- S. R. Fayers, N. H. Trewin, Trans. R. Soc. Edinburgh Earth Sci. 93, 355 (2003).
- M. W. Gaunt, M. A. Miles, Mol. Biol. Evol. 19, 748 (2002).
- D. Walossek, Fossils Strata 32, 54 (1993).
- This work was supported by The Danish Natural Science Council, The Velux Foundation, The Carlsberg Foundation, and The Wellcome Trust.
But remember: evolution is just a fairy-tale. All this "research" is really just a vast Darwinian conspiracy. Intelligent Design Creationism explains this stuff so much better! Nonetheless, it's fascinating stuff; to think about how much sense this makes, and how well-supported the evidence is, that Devil sure is tricky, eh?
________________
Technorati tags: Evolution